跑步如何导致贫血

跑步通常被认为是对健康最好的事情之一,从很多方面来说确实如此。它可以帮助改善您的姿势,降低患心脏病和糖尿病的风险,并增强下半身肌肉。
跑步通常被认为是对健康最好的事情之一,从很多方面来说确实如此。它可以帮助改善您的姿势,降低患心脏病和糖尿病的风险,并增强下半身肌肉。

天气会影响我们的情绪和健康——以下是贫血症患者和缺铁患者应该了解的有关低温的知识。

对于为什么女性比男性更容易贫血有一个简单的解释 - 以下是您可以采取的措施。
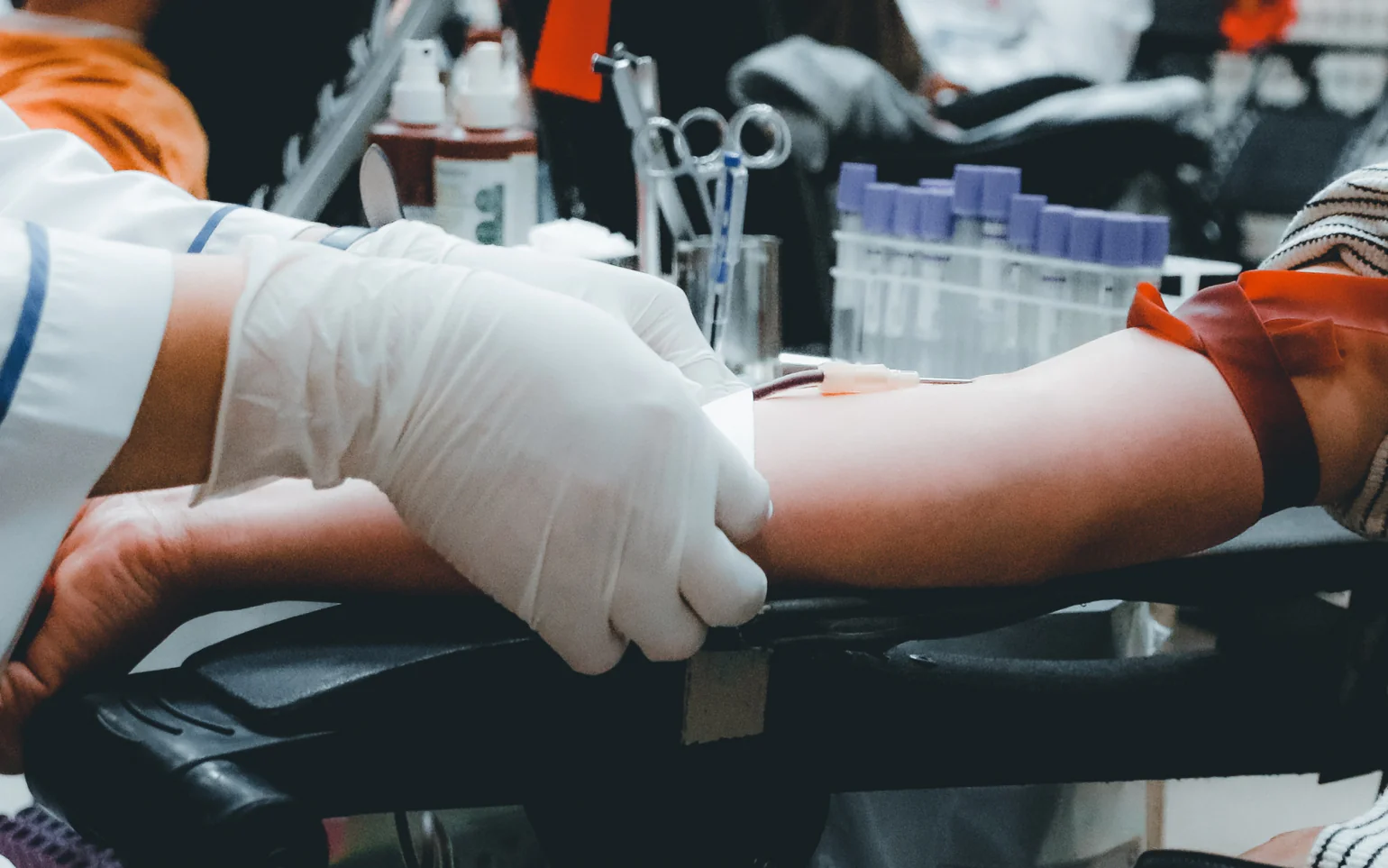
献血可以帮助那些接受择期手术或紧急手术的人,以及那些经常需要输血的人,例如重型 β 地中海贫血,每月需要输血四到五个献血者的血液。

如果您或您认识的人被诊断患有贫血,您可能会对它是什么以及它如何影响您的生活有一些疑问。贫血是一种影响全世界数百万人的疾病,了解它是获得所需治疗的第一步。让我们看一...

贫血症影响着全球超过 15 亿人,由于缺乏简单、负担得起的检测,其中许多人可能不知道自己的病情。它可能是一种难以忍受的疾病,甚至医生也很难正确识别和诊断患者。部分...

减肥可能具有挑战性,对于某些人来说,这似乎几乎是不可能的。然而,随着当前的减肥趋势,包括非处方减肥处方,越来越多的人开始转向这些快速解决方案。不幸的是,并非每种减...

您的预测中可能会出现缺铁性贫血——以下是您应该了解的内容。

炎症是身体自然愈合过程的重要组成部分。当身体的免疫系统对受伤、感染或任何其他导致组织损伤的触发因素做出反应时,就会发生这种情况。这是一种复杂的反应,涉及白细胞、细...

我们每天都被污染物包围——从我们呼吸的空气到我们消耗的食物,毒素几乎无处不在。不幸的是,这些毒素对我们的免疫系统和整体健康构成巨大威胁。让我们仔细看看毒素如何影响...

您是否注意到冬天起床变得更加困难?或者为什么不管前一天晚上睡了多少,白天你都会感到更累?这种现象比你想象的更常见,并且可以用科学来解释。我们来看看为什么换季的时候...

人们偶尔感到疲倦或情绪低落是很常见的。然而,如果你正与持续的疲劳和情绪低落作斗争,这可能是更重要的事情的征兆。根据研究,贫血和抑郁症之间有很强的联系。如果您或您认...
从向我们的细胞输送氧气到清除废物,血液对于我们身体的正常运作至关重要。良好的血液循环对于最佳健康至关重要,有很多方法可以改善或维持健康的循环。让我们来看看为什么良...

对于儿童、狗或猫来说,在 Zoom 上进行计划外的客串并不罕见,但对于 Rob Mannino 的团队来说,这种中断往往是一种挽救生命的过程。
以此为借口去享受阳光和维生素 D您是否注意到有多少音乐家真正歌颂太阳?歌手在每一首歌曲中如何讲述阳光如何让他们感觉良好?好吧,事实证明这是有一些充分的理由的。阳光...

糖已成为现代社会最受欢迎的食物之一。无论是糖果、苏打水还是烘焙食品,糖都已成为美国饮食中的主食。然而,它也是最有害的物质之一,可能导致慢性炎症和一系列健康问题。在...

您的红细胞可能会感谢您冷静下来——所以在阅读本文之前请深吸一口气:
镰状细胞病是一种遗传性血液疾病,影响全世界数百万人。据美国疾病控制与预防中心称,镰状细胞病影响着美国约 10 万人,其中非裔美国人受影响尤为严重。这是一种使人衰弱...
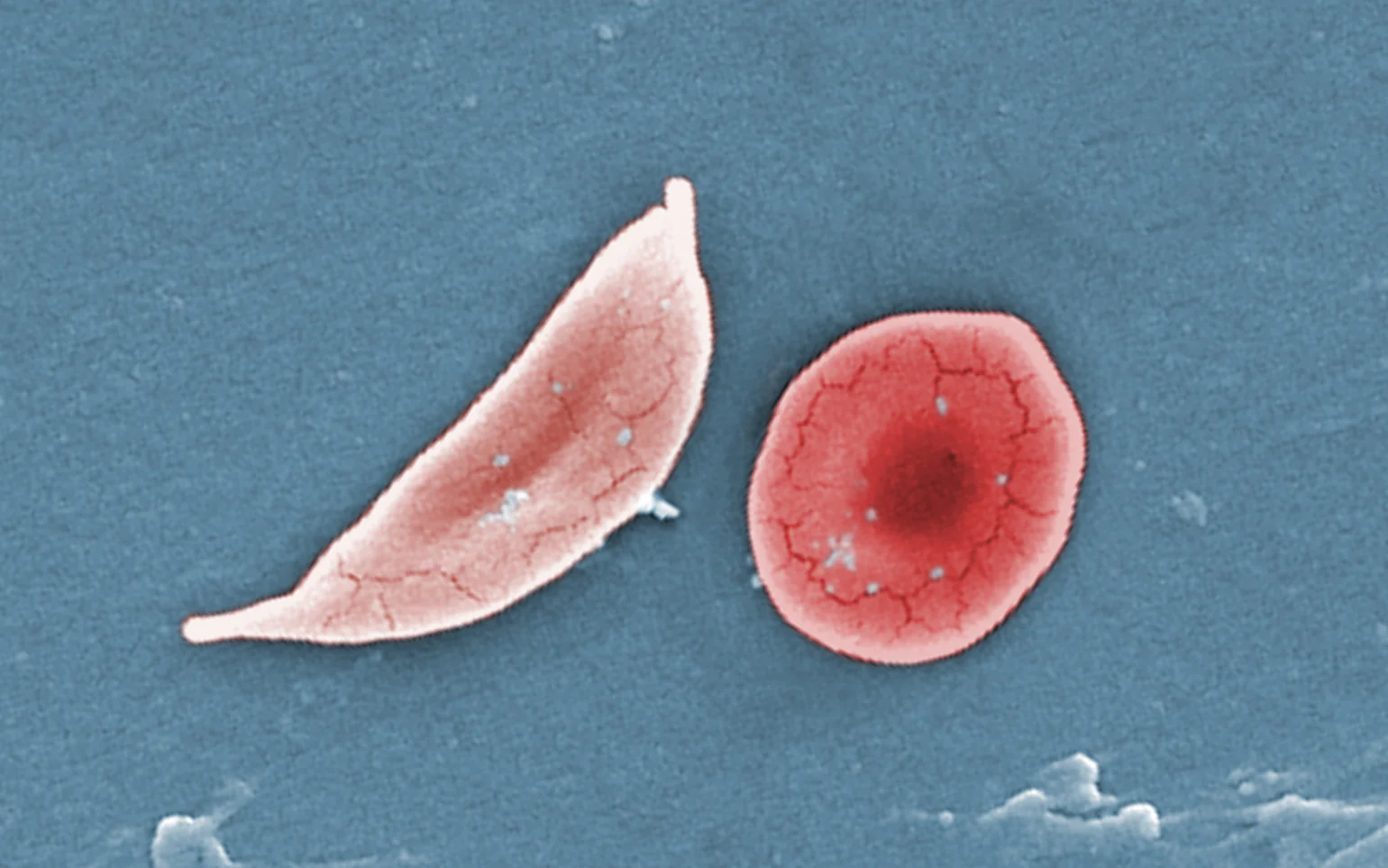
虽然这种疾病无法治愈,但进行一些教育可以大有帮助。

作为一名女性,花时间关注自己的健康并进行预防性护理非常重要。自检是您发现任何潜在问题的潜在迹象的最佳方法之一,如果不及早发现这些问题,这些问题可能会变得严重。
地中海贫血病也称为先天性溶血病。Vinmec医院的医生介绍,这是一种正常染色体上的隐性遗传病。而且男性和女性都可能发生。该疾病的特征是血红蛋白(红细胞结构中的重要...
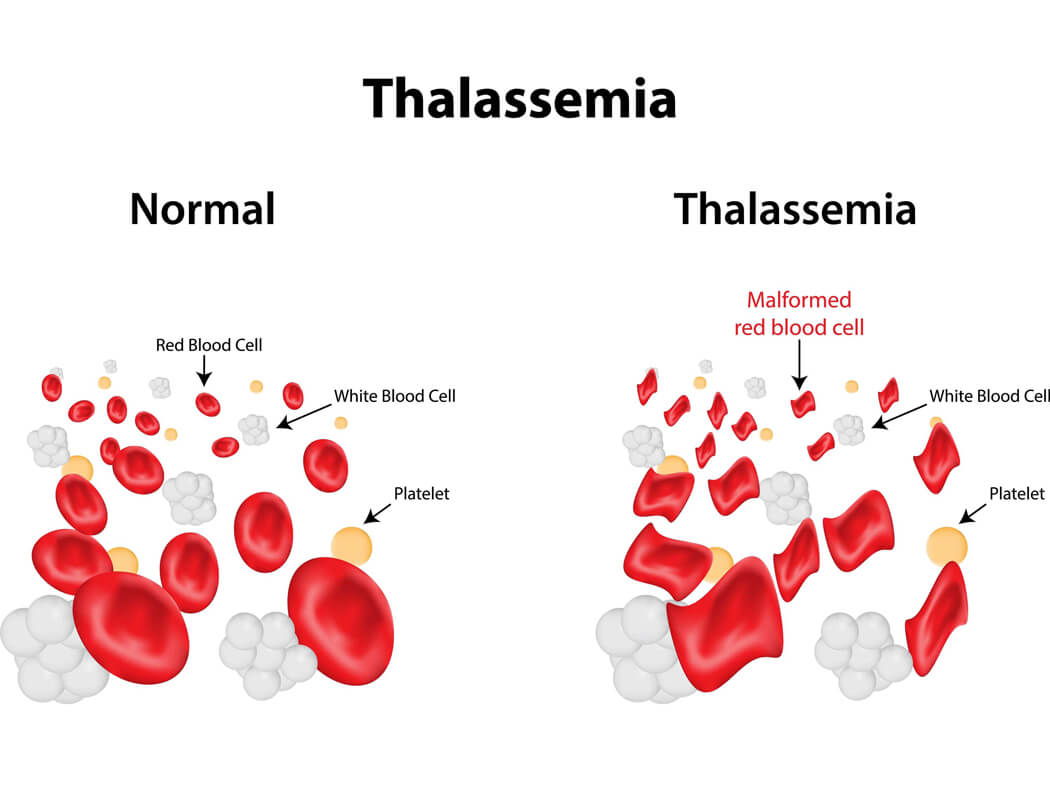
地中海贫血是一种遗传性疾病,会导致患者贫血和铁超载。那么地中海贫血的病因是什么呢?危险吗?以及如何治疗这种疾病呢?

地中海贫血是一种血液疾病,身体会产生异常形式的血红蛋白,血红蛋白是红细胞中携带氧气的蛋白质分子。这种疾病是由于红细胞过度破坏而导致的,导致贫血,即您的身体没有足够...

卵巢功能衰竭是一种直接影响女性整体健康及其生殖健康的疾病。很多女性担心卵巢功能衰竭能否治愈以及如何改善卵巢早衰。三英综合医院生殖支持中心的Ngo Dinh Tri...

拥有孩子是许多不孕夫妇的神圣愿望。胚胎移植到体内后,许多女性都会数着日子进行妊娠测试。那么胚胎移植后验孕棒要多久才能出结果呢?下面文章将为女性解答这个问题。

它是一种用于刺激卵泡成熟和刺激排卵的方法,用于治疗女性不孕症。HCG 注射在辅助生殖中非常受欢迎,有助于提高夫妇怀孕的成功率。什么是 hCG?人绒毛膜促性腺激素 ...

多囊卵巢综合症 (PCOS) 于 1935 年首次由两位美国妇科医生 Irving F Stein 和 Michael Leventhal 描述,这是女性中最常见...
地中海贫血是一种危险的遗传性疾病,在世界范围内很常见,估计有 7% 的人口携带该疾病基因。地中海贫血检测对于子孙后代治疗和预防该疾病至关重要。那么地中海贫血检查包...

化疗和放疗都会影响男性和女性的生育能力。如果您正在接受癌症治疗或已被诊断出患有癌症,您可能想知道未来是否要保留生育能力。
卵巢功能衰竭是一种危险的疾病,直接影响许多女性的整体健康以及生殖健康。尤其是在年轻人卵巢早衰呈上升趋势的背景下,很多女性都感到担忧,希望进行卵巢早衰检查,了解卵巢...
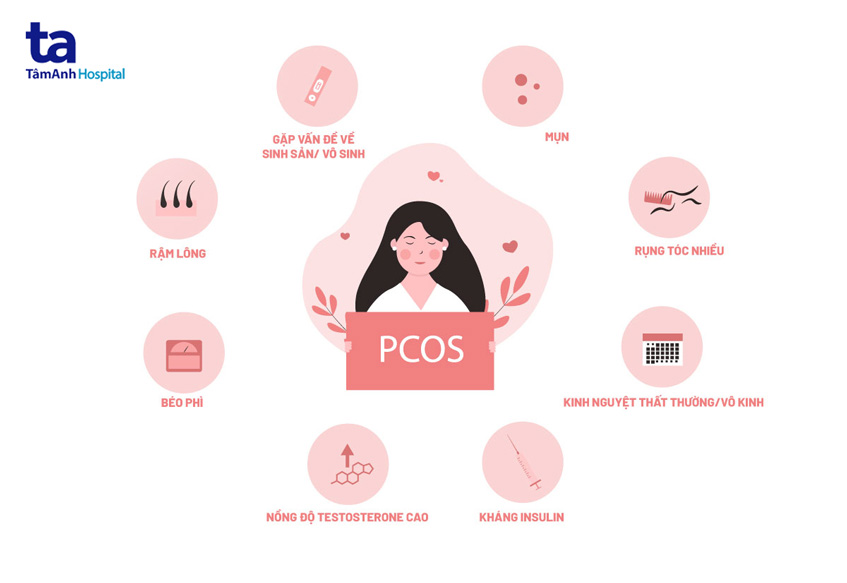
多囊卵巢综合症是女性非常常见的内分泌疾病。多囊卵巢综合症危害与否是很多女性的共同疑问。让我们在本文中了解该疾病可能出现的并发症。

输卵管阻塞是女性不孕的常见原因之一。每年约有 1000 万女性面临这种疾病。那么为什么输卵管堵塞会导致不孕呢?如何预防这种疾病,让我们在下面的文章中寻找答案。
如果您正在了解IVF ,您一定会听说过新鲜胚胎移植和冷冻胚胎移植的概念。那么,“为什么要移植冷冻胚胎?”“什么情况下应该移植?”“冷冻胚胎需要多少钱?”“冷冻胚胎...
地中海贫血(Thal)是一种遗传性溶血性贫血或先天性溶血性贫血,通过基因从父母遗传给子女,以贫血为主要症状。然而,可以通过父母的婚前检查、体外受精期间的胚胎活检或...

地中海贫血是一种遗传性(即通过基因从父母传给孩子)血液疾病,是由于身体无法产生足够的血红蛋白(红细胞的重要组成部分)蛋白质而引起的。当血红蛋白不足时,体内的红细胞...